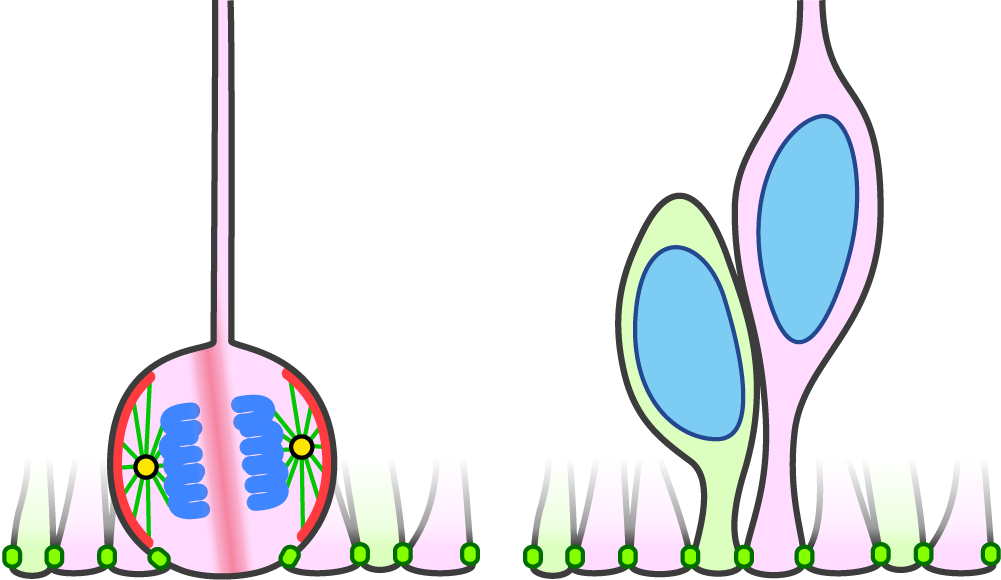
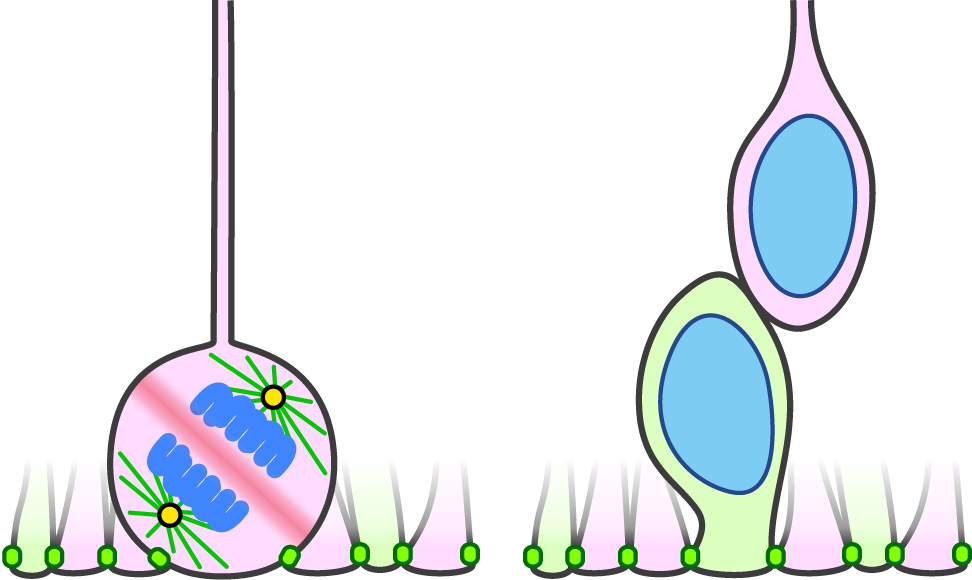
Neural stem cells (also known as radial glia, RG) maintain the epithelial structure during the mammalian cortical development. The spindle orientations in RG divisions are nearly planar to the ventricular surface, and thus the cleavage plane mostly bisects the apical surface, enabling both daughter cells to inherit the apical structure (Figure 1). The spindle orientation of RG is regulated by a dynein-anchoring complex containing LGN. The prevailing view claims that at the early proliferative stage, symmetric divisions of RG require tight regulation of the spindle orientation, the perturbation of which causes precocious neurogenesis and apoptosis. By contrast, previous studies indicated that at the subsequent neurogenic stage, spindle misorientations via LGN disruption mainly affect the migratory state and position of RG, rather than the daughter cell fates [1, 2]. LGN disruption in mice causes the cleavage plane to frequently bypass the apical surface, which results in the loss of the apical endfoot in one daughter cell. These daughter cells translocate to the basal side while maintaining the self-renewability and become outer/basal RG (oRG), a hallmark of gyrencephalic development (Figure 2). Thus, at the neurogenic stage, spindle orientation is essential for maintaining the RG epithelial structure but not for daughter cell fates or survival. Why RG show such differential properties depending on the stage was unsolved.
Figure 1. Spindle orientation of RG is planar to the apical surface in WT, so that the cleavage plane bissects the apical membrane (left). As a result, two daughter cells inherit the apical endfoot (right). Planar spindle orientation is mediated by LGN (red), which anchors astral microtubules to the cell cortex.
Figure 2. In the LGN mutant, spindle orientation is randomized. When the cleavage plane does not bissect the apical membrane (left), only the apical daughter inherits the apical endfoot and the basal daugher does not (right). Such basal daghuter translocates to the basal side with keeping self-renewability and becomes oRG at the neurogenic stage.
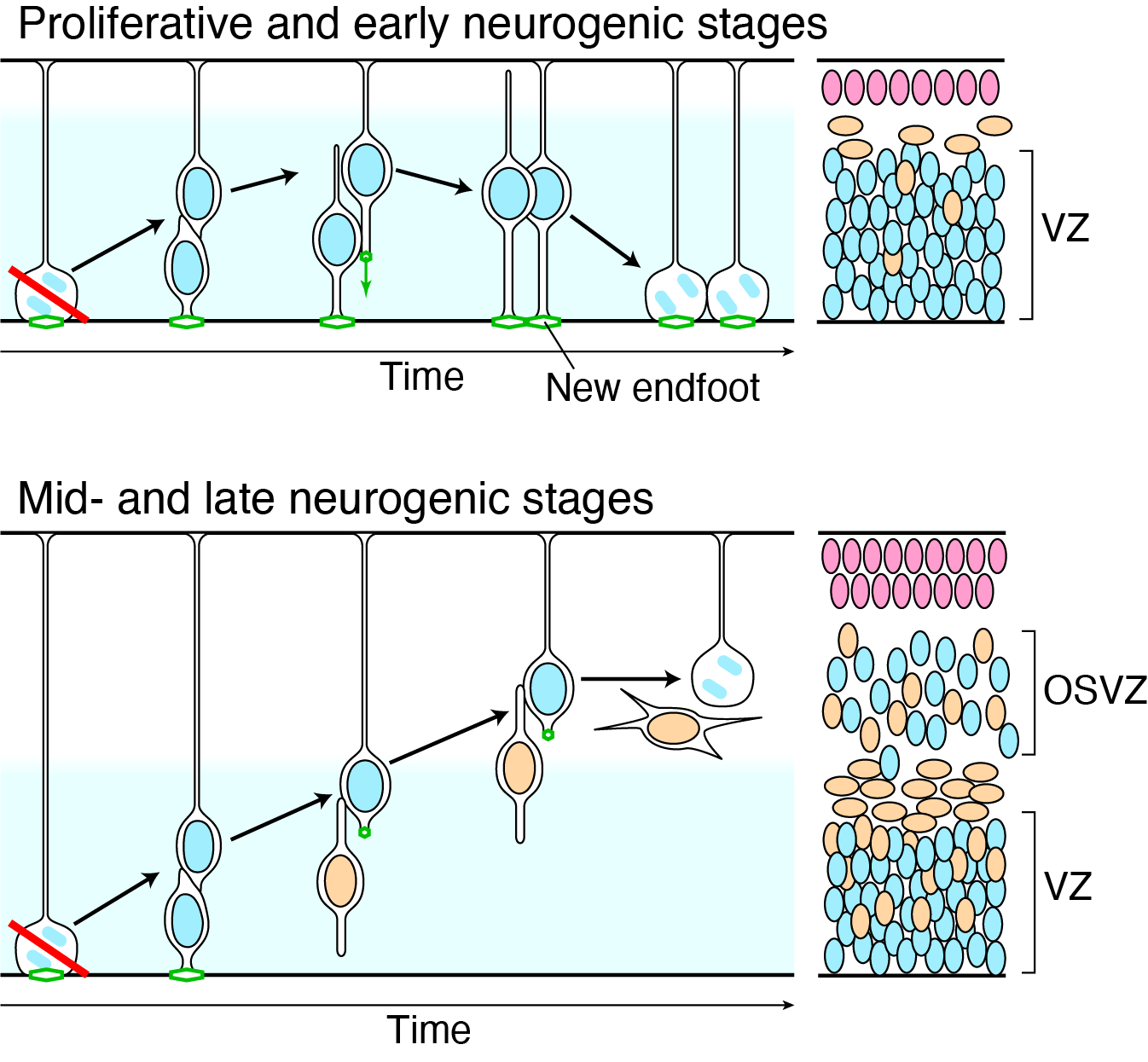
We showed that RG at the proliferative stage are highly robust against perturbation due to a regeneration ability of the epithelial structure but not a strict spindle orientation [3]. At the proliferative stage in the LGN mutant mice, spindle misorientations and consequent loss of the apical endfoot occurred as frequently as at the neurogenic stage. However, we observed neither precocious neurogenesis nor apoptosis at the proliferative stage. Instead, we found that the daughter cells missing the apical endfoot regenerated it, which led to recovery of the entire epithelial structure, and became RG. Regenerating endfeet attached to surrounding cells with ectopic adherens junctions at their leading edge. This ability gradually declined during neurogenesis, which allows RG missing the apical endfoot to transform into oRG at the late neurogenic stage (Figure 3). The regeneration ability was mediated by Notch–R-Ras–integrin β1 pathway, activity of which also declined during neurogenesis. Overactivation of this pathway in ferret, a gyrencephalic model animal, oRG formation was impaired.
Thus, our study revealed a temporally changing cryptic property, which initially ensures symmetric divisions of RG and subsequently provides a basis for the formation of the new germinal layer in the mammalian brain development.
Figure 3. Upper panel, during the early stage of the brain development, RG have the ability to regenerate the apical structure (green), so that RG can stay in the ventricular zone (VZ) and divide on the apical surface even if the spindle orientation is perturbed. Lower panel, loss of the regeneration ability leads to the generation of oRG and formation of outer subventricular zone (OSVZ) in the late stage of the brain development. Illustration: [3].
Microtubule-dependent nuclear oscillation
Introduction
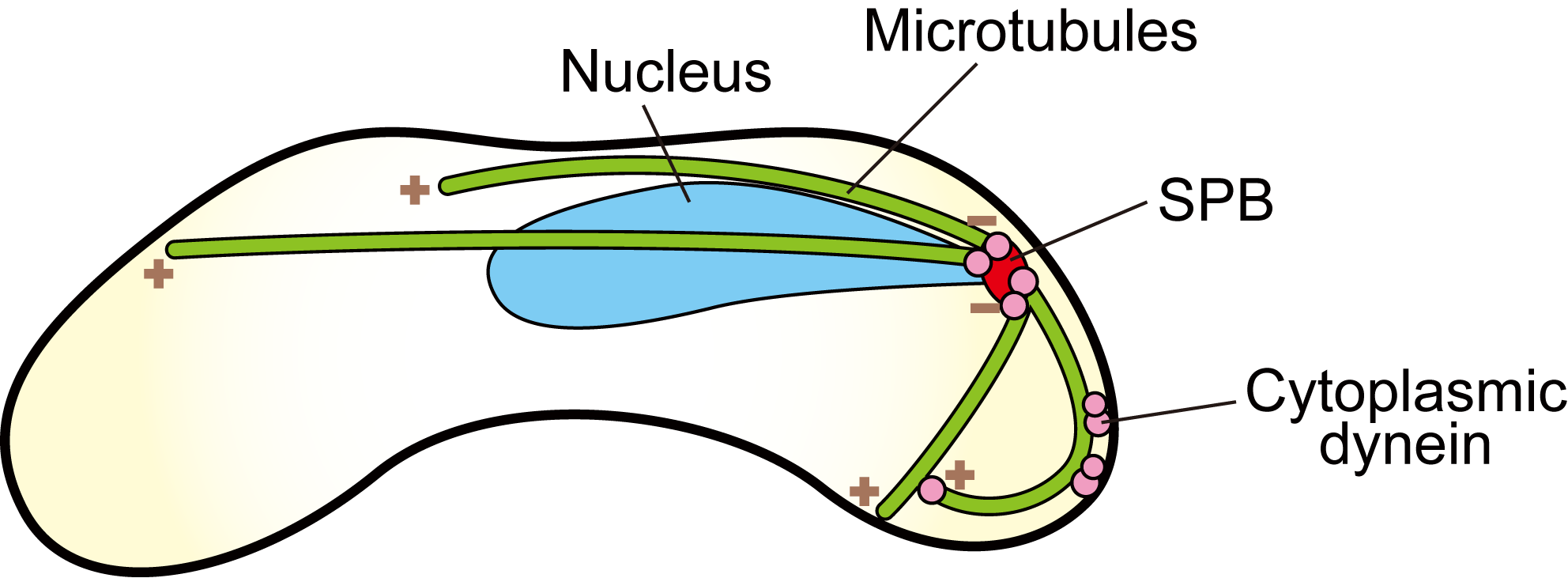
In fission yeast Schizosaccharomyces pombe, the nucleus led by the spindle pole body (SPB) oscillates between two poles of a cylindrical cell during meiotic S-phase to prophase. This nuclear oscillation is called “horsetail movement”. During the movement, all minus ends of microtubules are clustered at the SPB. A microtubule motor cytoplasmic dynein and dynactin are anchored to the cell cortex, of which movement toward the minus end of microtubules generates pulling force (Figure 4).
Figure 4. Schematic illustration of a fission yeast zygotic cell undergoing the horsetail movement. Green lines indicate microtubules, of which “+” and “−” represent the plus and minus ends, respectively. Cytoplasmic dynein anchored to the cell cortex moves toward the minus ends and generates pulling force on the SPB.
Dynein-anchoring mechanism
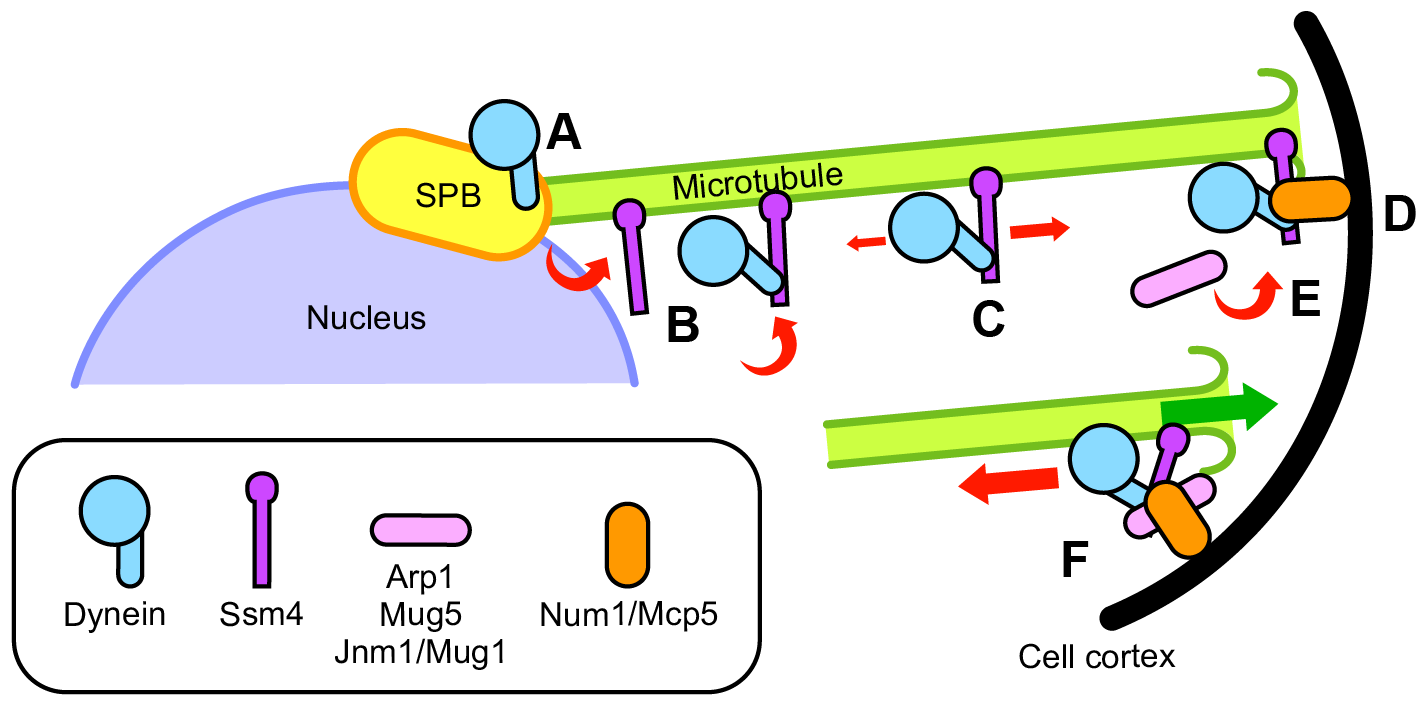
We identified three subunits of dynactin in fission yeast: Arp1, Mug5 and Jnm1/Mug1, and analyzed their functions in the horsetail movement. These subunits transiently colocalized with dynein foci at the cell cortex and were essential for the cortical anchoring of dynein. Cortical factor Num1/Mcp5, which was also required for dynein anchoring, bound to dynein independently of dynactin. Whereas Num1 suppressed the sliding of dynein foci along the cortex, Arp1, Mug5 and Jnm1 were involved in the regulation of shrinkage and bundling of microtubules. We proposed that dynein anchoring is established by cooperation of transient assembly of dynactin and Num1 at the cell cortex (Figure 5) [4, 5].
Figure 5. Model of cortical anchoring of cytoplasmic dynein in the horsetail movement. A, Dynein localizes to the SPB independently of dynactin. B, Dynein attaches to microtubules originating from the cytoplasm or being released from the SPB. C, Ssm4/p150Glued facilitates efficient binding of dynein to microtubules and allows dynein to disperse along microtubules, biasing towards plus ends. D, Dynein accumulates at shrinking plus ends and is captured by Num1/Mcp5 at the cell cortex. E, Arp1, Mug5/p24, and Jnm1/Mug1/p50/dynamitin assemble with the dynein–Ssm4 complex at the cell cortex and activate dynein. F, Assembly of dynactin and Num1 establishes stable anchoring of dynein to the cell cortex, enabling dynein to generate a microtubule-pulling force. Illustration: [5].
Oscillation mechanism
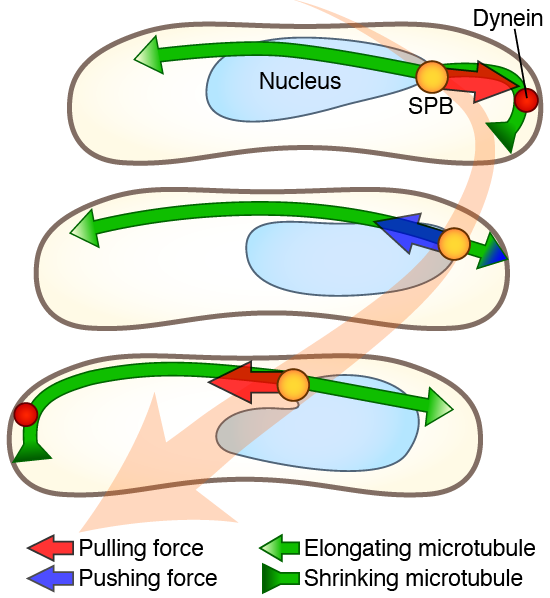
We studied how a robust oscillation is generated despite the dynamic behaviors of microtubules. We showed that the oscillation exhibits cell length-dependent frequency and requires a balance between microtubule and viscous drag forces, as well as proper microtubule dynamics (Figure 6) [6].
Comparison of the oscillations observed in living cells with a simulation model based on microtubule dynamic instability revealed that the period of oscillation correlates with cell length. Genetic alterations that reduce cargo size suggested that the nuclear movement depends on viscous drag forces. Deletion of a gene encoding Kinesin-8 inhibited microtubule catastrophe at the cell cortex and resulted in perturbation of oscillation, indicating that nuclear movement also depends on microtubule dynamic instability.
These findings linked numerical parameters from the simulation model with cellular functions required for generating the oscillation and provided a basis for understanding the physical properties of microtubule-dependent nuclear movements.
Figure 6. An oscillatory movement of the SPB (yellow circle) and the nucleus (sky blue) is driven by a coordination of the pulling and pushing forces on shrinking (inward green arrow) and growing (outward green arrow) microtubules. The red arrow indicates the pulling force generated by cortical cytoplasmic dynein (red circle). The blue arrow indicates the pushing force generated by short microtubules contacting the cell cortex. Illustration: [6].
References
Konno, D. et al. Neuroepithelial progenitors undergo LGN-dependent planar divisions to maintain self-renewability during mammalian neurogenesis. Nat. Cell Biol. 10, 93–101 (2008). DOI:10.1038/ncb1673.
Shitamukai, A., et al. Oblique radial glial divisions in the developing mouse neocortex induce self-renewing progenitors outside the germinal zone that resemble primate outer subventricular zone progenitors. J. Neurosci. 31, 3683–3695 (2011). DOI:10.1523/JNEUROSCI.4773-10.2011.
Fujita, I., et al. Endfoot regeneration restricts radial glial state and prevents translocation into the outer subventricular zone in early mammalian brain development. Nat. Cell Biol. 22, 26–37 (2020). DOI:10.1038/s41556-019-0436-9.
Fujita, I., et al. Contribution of dynein light intermediate and intermediate chains to subcellular localization of the dynein-dynactin motor complex in Schizosaccharomyces pombe. Genes Cells 15, 359–372 (2010). DOI:10.1111/j.1365-2443.2010.01386.x.
Fujita, I., et al. Dynactin and Num1 cooperate to establish the cortical anchoring of cytoplasmic dynein in S. pombe. J. Cell Sci. 128, 1555–1567 (2015). DOI:10.1242/jcs.163840.
Fujita, I., et al. A force balance model for a cell size-dependent meiotic nuclear oscillation in fission yeast. EMBO rep. 24, e55770 (2023). DOI:10.15252/embr.202255770.